
ma·lar·i·a (m -lâr
-lâr
 –)
–)
The advent of malaria must have been pretty appalling. Imagine a population with absolutely no resistance to the disease. Even today, after generations of resistance selected into Africans, malaria is one of the biggest killers there is, particularly of young children, killing about 1 million people a year in Africa. In a population with no resistance, it must have wiped out whole early farming communities in lowland areas.
In a remarkably short space of time, in evolutionary terms at least, there have been multiple responses selected into the African and other tropical populations. One of these, the Duffy negative allele, is now very common; over 95% of Sub Saharan Africans are Duffy negative. However, virtually all non Africans are Duffy positive, probably meaning that in the few thousand years that malaria has existed, natural selection scrubbed out all the D+ people in Africa, and rapidly replaced them with D- carriers (it’s a recessive gene too). My money goes on the early Bantu people as the origin for the D- mutation. The few exceptions to this are other malaria-prone areas like Borneo and the Andaman islands, who have independently had similar mutations to make them Duffy negative.
Some other blood adaptations to malaria.

More detailed information here.
This makes a total of at least fourteen evolutionary responses (not counting blood type O) to malaria in the space of about 5,000 years. The most notorious and second most common defence is sickle cell, which offers a protective effect if you only carry one gene, but gives you sickle cell disease if you carry both. The distribution of sickle cell very neatly matches the areas with the highest risk of malaria.


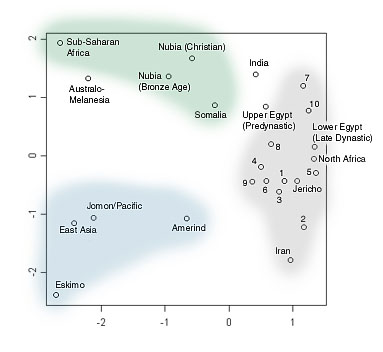
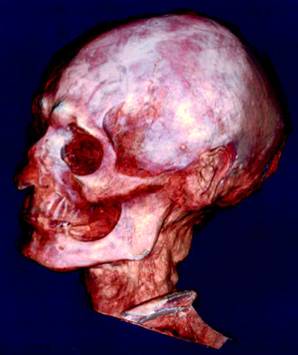
Some anthropologist once wrote that if you could magically transport a Cro Magnon human from 20,000 years ago into the 21st century, he’d die from the first bout of normal ‘flu he caught. Ever since humans started farming we’ve been exposed to an onslaught of infectious diseases that hunter gatherers never had to deal with. From the diseases caught from our animals (poxes and influenza) to normal human viruses given more virulence by increased population giving more scope for dangerous strains to arise, and more victims to catch them. These repeated selected sweeps for disease resistance are probably why the human skull shapes have seen more rapid divergence in the last 15,000 years than before (advent of farming).
I can’t help but wonder if the speed of Bantu expansion is partly due to their resistance to malaria. About 5,000 years ago the average African looked like a Khoisan, now they are mostly descended from Bantu settlers. Hunter gatherers generally don’t suffer as much from malaria, their population density is lower making the odds of an infected mosquito finding a human lower. But if you get a farming community turn up next door then there will be infected mosquitoes all over the place, and hunter gatherers just won’t have the same kind of resistance as the farmers. This would mean enough of the hunter gatherers would have died so the farmers have would find less opposition when they expand their territory into where the hunters were previously.





































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}